30.09.2019
Hvad hedder hullerne i cellemembranen? Membraner: deres struktur og funktion
Alle levende organismer, afhængigt af cellens struktur, er opdelt i tre grupper (se fig. 1):
1. Prokaryoter (ikke-nukleare)
2. Eukaryoter (nukleare)
3. Virus (ikke-cellulære)
Ris. 1. Levende organismer
I denne lektion vil vi begynde at studere strukturen af cellerne i eukaryote organismer, som omfatter planter, svampe og dyr. Deres celler er de største og mere komplekse sammenlignet med prokaryote celler.
Som du ved, er celler i stand til uafhængig aktivitet. De kan udveksle stof og energi med omgivelserne, samt vokse og formere sig, så cellens indre struktur er meget kompleks og primært afhængig af den funktion, som cellen udfører i en flercellet organisme.
Principperne for konstruktion af alle celler er de samme. I hver eukaryot celle kan følgende hoveddele skelnes (se fig. 2):
1. Den ydre membran, der adskiller cellens indhold fra det ydre miljø.
2. Cytoplasma med organeller.

Ris. 2. Hoveddelene af en eukaryot celle
Udtrykket "membran" blev foreslået for omkring hundrede år siden for at betegne cellens grænser, men med udviklingen af elektronmikroskopi blev det klart, at cellemembranen er en del af cellens strukturelle elementer.
I 1959 formulerede J. D. Robertson den elementære membranhypotese, hvorefter dyrs og planters cellemembraner er bygget efter samme type.
I 1972 blev det foreslået af Singer og Nicholson, hvilket i øjeblikket er almindeligt accepteret. Ifølge denne model er grundlaget for enhver membran et dobbeltlag af fosfolipider.
I fosfolipider (forbindelser indeholdende en fosfatgruppe) består molekylerne af et polært hoved og to ikke-polære haler (se fig. 3).

Ris. 3. Fosfolipid
I phospholipid-dobbeltlaget vender hydrofobe fedtsyrerester indad, mens hydrofile hoveder, inklusive en phosphorsyrerest, vender udad (se fig. 4).

Ris. 4. Fosfolipid-dobbeltlag
Fosfolipid-dobbeltlaget præsenteres som en dynamisk struktur, lipider kan bevæge sig og ændre deres position.
Det dobbelte lag af lipider giver membranens barrierefunktion, forhindrer cellens indhold i at sprede sig og forhindrer indtrængen af giftige stoffer i cellen.
Tilstedeværelsen af en grænsemembran mellem cellen og miljøet var kendt længe før fremkomsten af elektronmikroskopet. Fysiske kemikere benægtede eksistensen af plasmamembranen og mente, at der var en grænseflade mellem det levende kolloide indhold og miljøet, men Pfeffer (en tysk botaniker og plantefysiolog) i 1890 bekræftede dens eksistens.
I begyndelsen af forrige århundrede opdagede Overton (en britisk fysiolog og biolog) at penetrationshastigheden af mange stoffer i erytrocytter er direkte proportional med deres lipidopløselighed. I denne forbindelse foreslog forskeren, at membranen indeholder en stor mængde lipider og stoffer, der opløses i den, passerer gennem den og befinder sig på den anden side af membranen.
I 1925 isolerede Gorter og Grendel (amerikanske biologer) lipider fra erytrocytternes cellemembran. De resulterende lipider blev fordelt over vandoverfladen med en tykkelse på et molekyle. Det viste sig, at overfladearealet optaget af lipidlaget er to gange arealet af selve erytrocytten. Derfor konkluderede disse videnskabsmænd, at cellemembranen ikke består af et, men to lag af lipider.
Dawson og Danielli (engelske biologer) i 1935 foreslog, at det bimolekylære lipidlag i cellemembraner er indesluttet mellem to lag af proteinmolekyler (se fig. 5).

Ris. 5. Membranmodel foreslået af Dawson og Danielli
Med fremkomsten af elektronmikroskopet blev det muligt at sætte sig ind i membranens opbygning, og så fandt man ud af, at dyre- og plantecellers membraner ligner en trelagsstruktur (se fig. 6).

Ris. 6. Cellemembran under et mikroskop
I 1959 fremsatte biologen J. D. Robertson, ved at kombinere de data, der var tilgængelige på det tidspunkt, en hypotese om strukturen af den "elementære membran", hvori han postulerede en struktur, der er fælles for alle biologiske membraner.
Robertsons postulater om strukturen af den "elementære membran"
1. Alle membraner er omkring 7,5 nm tykke.
2. I et elektronmikroskop ser de alle ud til at være trelagede.
3. Trelagsbilledet af membranen er resultatet af nøjagtigt arrangementet af proteiner og polære lipider, som blev leveret af Dawson og Danielli-modellen - det centrale lipiddobbeltlag er indesluttet mellem to lag protein.
Denne hypotese om strukturen af den "elementære membran" har undergået forskellige ændringer, og i 1972 blev den fremsat af flydende mosaikmodel af membranen(se fig. 7), hvilket nu er almindeligt accepteret.

Ris. 7. Flydende mosaikmodel af membranen
Proteinmolekyler er nedsænket i lipid-dobbeltlaget af membranen, de danner en mobil mosaik. I henhold til deres placering i membranen og den måde, de interagerer med lipiddobbeltlaget, kan proteiner opdeles i:
- overfladisk (eller perifer) membranproteiner forbundet med den hydrofile overflade af lipiddobbeltlaget;
- integral (membran) proteiner indlejret i det hydrofobe område af dobbeltlaget.
Integrale proteiner adskiller sig i graden af deres nedsænkning i det hydrofobe område af dobbeltlaget. De kan være helt nedsænket integral) eller delvist nedsænket ( semi-integral), og kan også trænge ind i membranen gennem ( transmembran).
Membranproteiner kan opdeles i to grupper efter deres funktioner:
- strukturel proteiner. De er en del af cellemembraner og er involveret i at opretholde deres struktur.
- dynamisk proteiner. De er placeret på membranerne og deltager i de processer, der finder sted på den.
Der er tre klasser af dynamiske proteiner.
1. Receptor. Ved hjælp af disse proteiner opfatter cellen forskellige påvirkninger på dens overflade. Det vil sige, at de specifikt binder forbindelser som hormoner, neurotransmittere, toksiner på ydersiden af membranen, hvilket fungerer som et signal til at ændre forskellige processer inde i cellen eller selve membranen.
2. Transportere. Disse proteiner transporterer visse stoffer gennem membranen, de danner også kanaler, hvorigennem forskellige ioner transporteres ind og ud af cellen.
3. Enzymatisk. Det er enzymproteiner, der er placeret i membranen og er involveret i forskellige kemiske processer.
Transport af stoffer over membranen
Lipid-dobbeltlag er stort set uigennemtrængelige for mange stoffer, så der kræves en stor mængde energi for at transportere stoffer gennem membranen, og der kræves også dannelse af forskellige strukturer.
Der er to typer transport: passiv og aktiv.
Passiv transport
Passiv transport er bevægelsen af molekyler langs en koncentrationsgradient. Det vil sige, at det kun bestemmes af forskellen i koncentrationen af det overførte stof på modsatte sider af membranen og udføres uden energiforbrug.
Der er to typer passiv transport:
- simpel diffusion(se fig. 8), hvilket sker uden deltagelse af membranproteinet. Mekanismen for simpel diffusion er transmembranoverførsel af gasser (ilt og kuldioxid), vand og nogle simple organiske ioner. Simpel diffusion er langsom.

Ris. 8. Simpel diffusion
- lettet diffusion(se fig. 9) adskiller sig fra simple ved, at det foregår med deltagelse af bærerproteiner. Denne proces er specifik og forløber med en højere hastighed end simpel diffusion.

Ris. 9. Faciliteret diffusion
Der kendes to typer membrantransportproteiner: bærerproteiner (translokaser) og kanaldannende proteiner. Transportproteiner binder specifikke stoffer og fører dem over membranen langs deres koncentrationsgradient, og derfor kræver denne proces, som ved simpel diffusion, ikke forbrug af ATP-energi.
Fødepartikler kan ikke passere gennem membranen, de kommer ind i cellen ved endocytose (se fig. 10). Under endocytose danner plasmamembranen invaginationer og udvækster, fanger en fast fødepartikel. En vakuole (eller vesikel) dannes omkring madbolusen, som så løsnes fra plasmamembranen, og den faste partikel i vakuolen er inde i cellen.

Ris. 10. Endocytose
Der er to typer endocytose.
1. Fagocytose- absorption af faste partikler. Specialiserede celler, der udfører fagocytose kaldes fagocytter.
2. pinocytose- absorption af flydende materiale (opløsning, kolloid opløsning, suspensioner).
Exocytose(se fig. 11) - en proces omvendt til endocytose. Stoffer, der syntetiseres i cellen, såsom hormoner, pakkes ind i membranvesikler, der passer til cellemembranen, indlejres i den, og vesiklens indhold udstødes fra cellen. På samme måde kan cellen komme af med unødvendige stofskifteprodukter.

Ris. 11. Exocytose
aktiv transport
I modsætning til faciliteret diffusion er aktiv transport bevægelse af stoffer mod en koncentrationsgradient. I dette tilfælde flytter stoffer sig fra et område med en lavere koncentration til et område med en højere koncentration. Da en sådan bevægelse sker i den modsatte retning af normal diffusion, skal cellen bruge energi i denne proces.
Blandt eksempler på aktiv transport studeres bedst den såkaldte natrium-kalium-pumpe. Denne pumpe pumper natriumioner ud af cellen og pumper kaliumioner ind i cellen ved hjælp af energien fra ATP.
1. Strukturel (cellemembranen adskiller cellen fra miljøet).
2. Transport (stoffer transporteres gennem cellemembranen, og cellemembranen er et meget selektivt filter).
3. Receptor (receptorer placeret på overfladen af membranen opfatter ydre påvirkninger, transmitterer denne information ind i cellen, så den hurtigt reagerer på miljøændringer).
Ud over de ovenfor nævnte udfører membranen også en metabolisk og energiomdannende funktion.
metabolisk funktion
Biologiske membraner er direkte eller indirekte involveret i processerne af metaboliske transformationer af stoffer i cellen, da de fleste enzymer er forbundet med membraner.
Lipidmiljøet af enzymer i membranen skaber visse betingelser for deres funktion, pålægger begrænsninger for aktiviteten af membranproteiner og har således en regulerende effekt på metaboliske processer.
Energikonverteringsfunktion
Den vigtigste funktion af mange biomembraner er omdannelsen af en form for energi til en anden.
Energikonverterende membraner omfatter indre membraner af mitokondrier, thylakoider af kloroplaster (se fig. 12).

Ris. 12. Mitokondrier og kloroplast
Bibliografi
- Kamensky A.A., Kriksunov E.A., Pasechnik V.V. Generel biologi 10-11 klasse Bustard, 2005.
- Biologi. 10. klasse. Generel biologi. Grundniveau / P.V. Izhevsky, O.A. Kornilova, T.E. Loshchilin og andre - 2. udg., revideret. - Ventana-Graf, 2010. - 224 sider.
- Belyaev D.K. Biologi 10-11 klasse. Generel biologi. Et grundlæggende niveau af. - 11. udgave, stereotype. - M.: Uddannelse, 2012. - 304 s.
- Agafonova I.B., Zakharova E.T., Sivoglazov V.I. Biologi 10-11 klasse. Generel biologi. Et grundlæggende niveau af. - 6. udg., tilføje. - Bustard, 2010. - 384 s.
- Ayzdorov.ru ().
- Youtube.com().
- Doctor-v.ru ().
- Animals-world.ru ().
Lektier
- Hvad er opbygningen af en cellemembran?
- Hvilke egenskaber har lipider til at danne membraner?
- På grund af hvilke funktioner er proteiner i stand til at deltage i transporten af stoffer over membranen?
- Angiv plasmamembranens funktioner.
- Hvordan foregår passiv transport over membranen?
- Hvordan foregår aktiv transport over membranen?
- Hvad er funktionen af natrium-kalium pumpen?
- Hvad er fagocytose, pinocytose?
Studiet af strukturen af organismer, såvel som planter, dyr og mennesker, er den gren af biologien kaldet cytologi. Forskere har fundet ud af, at indholdet af cellen, som er inde i den, er ret komplekst. Det er omgivet af det såkaldte overfladeapparat, som omfatter den ydre cellemembran, supra-membranstrukturer: glycocalyx og mikrofilamenter, pelicule og mikrotubuli, der danner dens submembrankompleks.
I denne artikel vil vi studere strukturen og funktionerne af den ydre cellemembran, som er en del af overfladeapparatet på forskellige celletyper.
Hvad er den ydre cellemembrans funktioner?
Som beskrevet tidligere er den ydre membran en del af overfladeapparatet i hver celle, som med succes adskiller dets indre indhold og beskytter celleorganeller mod ugunstige miljøforhold. En anden funktion er at sikre udvekslingen af stoffer mellem celleindholdet og vævsvæsken, derfor transporterer den ydre cellemembran molekyler og ioner ind i cytoplasmaet, og hjælper også med at fjerne toksiner og overskydende giftige stoffer fra cellen.

Strukturen af cellemembranen
Membraner, eller plasmalemmaer, af forskellige typer celler er meget forskellige fra hinanden. Hovedsageligt den kemiske struktur, såvel som det relative indhold af lipider, glycoproteiner, proteiner i dem og følgelig arten af receptorerne i dem. Ekstern, som primært bestemmes af den individuelle sammensætning af glycoproteiner, deltager i genkendelsen af miljøstimuli og i selve cellens reaktioner på deres handlinger. Nogle typer vira kan interagere med proteiner og glykolipider i cellemembraner, som et resultat af hvilke de trænger ind i cellen. Herpes og influenzavirus kan bruge til at bygge deres beskyttende skal.
Og vira og bakterier, de såkaldte bakteriofager, binder sig til cellemembranen og opløser den ved kontaktpunktet ved hjælp af et særligt enzym. Derefter passerer et molekyle af viralt DNA ind i det dannede hul.
Funktioner af strukturen af plasmamembranen af eukaryoter
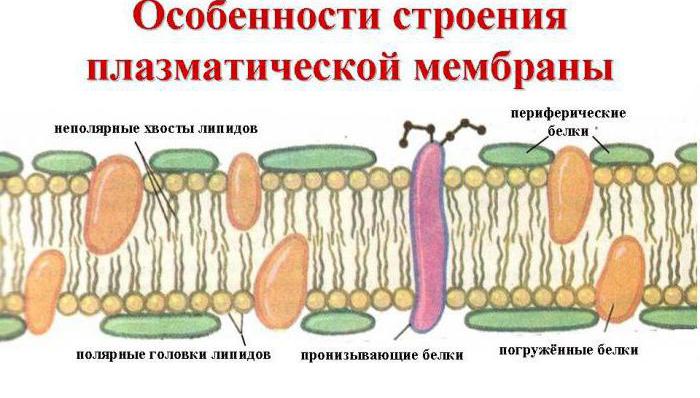
Husk på, at den ydre cellemembran udfører transportfunktionen, det vil sige overførsel af stoffer ind i og ud af den i det ydre miljø. For at udføre en sådan proces kræves en speciel struktur. Faktisk er plasmalemmaet et konstant, universelt system af overfladeapparatet for alle. Dette er en tynd (2-10 Nm), men ret tæt flerlagsfilm, der dækker hele cellen. Dens struktur blev undersøgt i 1972 af sådanne videnskabsmænd som D. Singer og G. Nicholson, de skabte også en væske-mosaikmodel af cellemembranen.
De vigtigste kemiske forbindelser, der danner det, er ordnede molekyler af proteiner og visse fosfolipider, som er spredt i et flydende lipidmiljø og ligner en mosaik. Således består cellemembranen af to lag af lipider, hvis ikke-polære hydrofobe "haler" er inde i membranen, og de polære hydrofile hoveder vender mod cellens cytoplasma og den interstitielle væske.
Lipidlaget er gennemtrængt af store proteinmolekyler, der danner hydrofile porer. Det er gennem dem, at vandige opløsninger af glucose og mineralsalte transporteres. Nogle proteinmolekyler er placeret både på den ydre og indre overflade af plasmalemmaet. På den ydre cellemembran i cellerne i alle organismer med kerner er der således kulhydratmolekyler bundet af kovalente bindinger med glykolipider og glykoproteiner. Indholdet af kulhydrater i cellemembraner varierer fra 2 til 10 %.

Strukturen af plasmalemmaet af prokaryote organismer
Den ydre cellemembran i prokaryoter udfører lignende funktioner som plasmamembranerne i celler fra kerneorganismer, nemlig: perception og transmission af information, der kommer fra det ydre miljø, transport af ioner og opløsninger ind og ud af cellen, og beskyttelse af cytoplasmaet fra fremmede reagenser udefra. Det kan danne mesosomer – strukturer, der opstår, når plasmalemmaet stikker ind i cellen. De kan indeholde enzymer involveret i de metaboliske reaktioner af prokaryoter, for eksempel i DNA-replikation, proteinsyntese.
Mesosomer indeholder også redoxenzymer, mens fotosyntetika indeholder bakteriochlorophyll (i bakterier) og phycobilin (i cyanobakterier).
Ydre membraners rolle i intercellulære kontakter
Lad os fortsætte med at besvare spørgsmålet om, hvilke funktioner den ydre cellemembran udfører, så lad os dvæle ved dens rolle i planteceller.I planteceller dannes porer i væggene i den ydre cellemembran, der passerer ind i celluloselaget. Gennem dem er udgangen af cellens cytoplasma til ydersiden mulig; sådanne tynde kanaler kaldes plasmodesmata.

Takket være dem er forbindelsen mellem naboplanteceller meget stærk. I menneske- og dyreceller kaldes kontaktstederne mellem tilstødende cellemembraner desmosomer. De er karakteristiske for endotel- og epitelceller og findes også i kardiomyocytter.
Hjælpeformationer af plasmalemmaet
For at forstå, hvordan planteceller adskiller sig fra dyr, hjælper det at studere de strukturelle træk ved deres plasmamembraner, som afhænger af, hvilke funktioner den ydre cellemembran udfører. Over det i dyreceller er et lag af glycocalyx. Det er dannet af polysaccharidmolekyler forbundet med proteiner og lipider i den ydre cellemembran. Takket være glycocalyxen opstår adhæsion (klæbning) mellem celler, hvilket fører til dannelsen af væv, derfor deltager det i plasmalemmaets signalfunktion - genkendelsen af miljøstimuli.
Hvordan er den passive transport af visse stoffer over cellemembraner
Som tidligere nævnt er den ydre cellemembran involveret i processen med at transportere stoffer mellem cellen og det ydre miljø. Der er to typer transport gennem plasmalemmaet: passiv (diffusion) og aktiv transport. Den første omfatter diffusion, faciliteret diffusion og osmose. Bevægelsen af stoffer langs koncentrationsgradienten afhænger primært af massen og størrelsen af de molekyler, der passerer gennem cellemembranen. For eksempel opløses små ikke-polære molekyler let i det midterste lipidlag af plasmalemmaet, bevæger sig igennem det og ender i cytoplasmaet.

Store molekyler af organiske stoffer trænger ind i cytoplasmaet ved hjælp af specielle bærerproteiner. De er artsspecifikke og, når de kombineres med en partikel eller ion, overfører de dem passivt gennem membranen langs koncentrationsgradienten uden at bruge energi (passiv transport). Denne proces ligger til grund for en sådan egenskab ved plasmalemmaet som selektiv permeabilitet. I processen bruges energien fra ATP-molekyler ikke, og cellen gemmer den til andre metaboliske reaktioner.
Aktiv transport af kemiske forbindelser over plasmalemmaet
Da den ydre cellemembran sikrer overførsel af molekyler og ioner fra det ydre miljø ind i cellen og tilbage, bliver det muligt at fjerne dissimileringsprodukterne, som er toksiner, til ydersiden, altså til den intercellulære væske. opstår mod en koncentrationsgradient og kræver brug af energi i form af ATP-molekyler. Det involverer også bærerproteiner kaldet ATPaser, som også er enzymer.

Et eksempel på sådan transport er natrium-kalium-pumpen (natriumioner passerer fra cytoplasmaet til det ydre miljø, og kaliumioner pumpes ind i cytoplasmaet). Tarmens og nyrernes epitelceller er i stand til det. Varianter af denne metode til overførsel er processerne af pinocytose og fagocytose. Efter at have studeret, hvilke funktioner den ydre cellemembran udfører, kan det fastslås, at heterotrofe protister såvel som celler fra højere dyreorganismer, for eksempel leukocytter, er i stand til pino- og fagocytose.
Bioelektriske processer i cellemembraner
Det er blevet fastslået, at der er en potentiel forskel mellem den ydre overflade af plasmalemmaet (det er positivt ladet) og det parietale lag af cytoplasmaet, som er negativt ladet. Det blev kaldt hvilepotentialet, og det er iboende i alle levende celler. Og nervevævet har ikke kun et hvilepotentiale, men er også i stand til at lede svage biostrømme, som kaldes excitationsprocessen. De ydre membraner af nerveceller-neuroner, der modtager irritation fra receptorer, begynder at ændre ladninger: natriumioner kommer massivt ind i cellen, og overfladen af plasmalemmaet bliver elektronegativ. Og det parietale lag af cytoplasmaet, på grund af et overskud af kationer, modtager en positiv ladning. Dette forklarer, hvorfor neurons ydre cellemembran genoplades, hvilket forårsager ledning af nerveimpulser, der ligger til grund for excitationsprocessen.
celle membran
Billede af en cellemembran. Små blå og hvide kugler svarer til de hydrofobe "hoveder" af fosfolipiderne, og linjerne knyttet til dem svarer til de hydrofile "haler". Figuren viser kun integrale membranproteiner (røde kugler og gule helixer). Gule ovale prikker inde i membranen - kolesterolmolekyler Gulgrønne kæder af perler på ydersiden af membranen - oligosaccharidkæder, der danner glykokalyxen
Den biologiske membran omfatter også forskellige proteiner: integral (penetrerer membranen igennem), semi-integral (nedsænket i den ene ende i det ydre eller indre lipidlag), overflade (placeret på den ydre eller støder op til de indvendige sider af membranen). Nogle proteiner er cellemembranens kontaktpunkter med cytoskelettet inde i cellen og cellevæggen (hvis nogen) udenfor. Nogle af de integrerede proteiner fungerer som ionkanaler, forskellige transportører og receptorer.
Funktioner
- barriere - giver et reguleret, selektivt, passivt og aktivt stofskifte med omgivelserne. For eksempel beskytter peroxisommembranen cytoplasmaet mod peroxider, der er farlige for cellen. Selektiv permeabilitet betyder, at en membrans permeabilitet for forskellige atomer eller molekyler afhænger af deres størrelse, elektriske ladning og kemiske egenskaber. Selektiv permeabilitet sikrer adskillelsen af cellen og cellulære rum fra miljøet og forsyner dem med de nødvendige stoffer.
- transport - gennem membranen sker der en transport af stoffer ind i cellen og ud af cellen. Transport gennem membranerne giver: tilførsel af næringsstoffer, fjernelse af slutprodukter af stofskiftet, udskillelse af forskellige stoffer, skabelse af ioniske gradienter, opretholdelse af det optimale og koncentration af ioner i cellen, som er nødvendige for at fungere af cellulære enzymer.
Partikler, der af en eller anden grund ikke er i stand til at krydse fosfolipid-dobbeltlaget (for eksempel på grund af hydrofile egenskaber, da membranen indeni er hydrofob og ikke tillader hydrofile stoffer at passere igennem, eller på grund af deres store størrelse), men nødvendige for cellen , kan trænge ind i membranen gennem specielle bærerproteiner (transportører) og kanalproteiner eller ved endocytose.
Ved passiv transport krydser stoffer lipid-dobbeltlaget uden energiforbrug langs koncentrationsgradienten ved diffusion. En variant af denne mekanisme er faciliteret diffusion, hvor et specifikt molekyle hjælper et stof med at passere gennem membranen. Dette molekyle kan have en kanal, der tillader kun én type stof at passere igennem.
Aktiv transport kræver energi, da den sker mod en koncentrationsgradient. Der er specielle pumpeproteiner på membranen, herunder ATPase, som aktivt pumper kaliumioner (K+) ind i cellen og pumper natriumioner (Na+) ud af den. - matrix - giver en vis relativ position og orientering af membranproteiner, deres optimale interaktion.
- mekanisk - sikrer cellens autonomi, dens intracellulære strukturer samt forbindelse med andre celler (i væv). Cellevægge spiller en vigtig rolle i at give mekanisk funktion, og hos dyr - intercellulært stof.
- energi - under fotosyntese i kloroplaster og cellulær respiration i mitokondrier fungerer energioverførselssystemer i deres membraner, hvori proteiner også deltager;
- receptor - nogle proteiner placeret i membranen er receptorer (molekyler, som cellen opfatter bestemte signaler med).
For eksempel virker hormoner, der cirkulerer i blodet, kun på målceller, der har receptorer, der svarer til disse hormoner. Neurotransmittere (kemikalier, der leder nerveimpulser) binder sig også til specifikke receptorproteiner på målceller. - enzymatiske - membranproteiner er ofte enzymer. For eksempel indeholder plasmamembranerne i tarmepitelcellerne fordøjelsesenzymer.
- implementering af generering og ledning af biopotentialer.
Ved hjælp af membranen opretholdes en konstant koncentration af ioner i cellen: koncentrationen af K + ionen inde i cellen er meget højere end udenfor, og koncentrationen af Na + er meget lavere, hvilket er meget vigtigt, da dette opretholder potentialeforskellen over membranen og genererer en nerveimpuls. - cellemarkering - der er antigener på membranen, der fungerer som markører - "mærker", der gør det muligt at identificere cellen. Disse er glykoproteiner (det vil sige proteiner med forgrenede oligosaccharid-sidekæder knyttet til dem), der spiller rollen som "antenner". På grund af de utallige sidekædekonfigurationer er det muligt at lave en specifik markør for hver celletype. Ved hjælp af markører kan celler genkende andre celler og handle sammen med dem, for eksempel ved dannelse af organer og væv. Det tillader også immunsystemet at genkende fremmede antigener.
Struktur og sammensætning af biomembraner
Membraner er sammensat af tre klasser af lipider: fosfolipider, glykolipider og kolesterol. Fosfolipider og glycolipider (lipider med kulhydrater knyttet til dem) består af to lange hydrofobe kulbrinte-"haler", der er forbundet med et ladet hydrofilt "hoved". Kolesterol stivner membranen ved at optage det frie rum mellem de hydrofobe lipidhaler og forhindre dem i at bøje. Derfor er membraner med et lavt kolesterolindhold mere fleksible, mens dem med et højt kolesterolindhold er mere stive og skøre. Kolesterol fungerer også som en "prop", der forhindrer bevægelse af polære molekyler fra og ind i cellen. En vigtig del af membranen består af proteiner, der trænger ind i den og er ansvarlige for forskellige egenskaber ved membraner. Deres sammensætning og orientering i forskellige membraner er forskellig.
Cellemembraner er ofte asymmetriske, det vil sige, at lagene adskiller sig i lipidsammensætning, overgangen af et individuelt molekyle fra et lag til et andet (den såkaldte klipklapper) er svært.
Membranorganeller
Disse er lukkede enkelte eller indbyrdes forbundne sektioner af cytoplasmaet, adskilt fra hyaloplasmaet af membraner. Enkeltmembranorganeller indbefatter endoplasmatisk retikulum, Golgi-apparater, lysosomer, vakuoler, peroxisomer; til to-membran - kerne, mitokondrier, plastider. Strukturen af membranerne i forskellige organeller adskiller sig i sammensætningen af lipider og membranproteiner.
Selektiv permeabilitet
Cellemembraner har selektiv permeabilitet: Glucose, aminosyrer, fedtsyrer, glycerol og ioner diffunderer langsomt gennem dem, og membranerne selv regulerer aktivt denne proces i et vist omfang - nogle stoffer passerer igennem, mens andre ikke gør det. Der er fire hovedmekanismer for indtrængen af stoffer i cellen eller deres fjernelse fra cellen til ydersiden: diffusion, osmose, aktiv transport og exo- eller endocytose. De to første processer er passive i naturen, det vil sige, at de ikke kræver energi; de sidste to er aktive processer forbundet med energiforbrug.
Membranens selektive permeabilitet under passiv transport skyldes specielle kanaler - integrerede proteiner. De trænger igennem og gennem membranen og danner en slags passage. Grundstofferne K, Na og Cl har deres egne kanaler. Med hensyn til koncentrationsgradienten bevæger molekylerne af disse grundstoffer sig ind og ud af cellen. Ved irritation åbner natriumionkanalerne sig, og der sker en skarp indstrømning af natriumioner ind i cellen. Dette resulterer i en ubalance i membranpotentialet. Derefter genoprettes membranpotentialet. Kaliumkanaler er altid åbne, hvorigennem kaliumioner langsomt kommer ind i cellen.
se også
Litteratur
- Antonov V. F., Smirnova E. N., Shevchenko E. V. Lipidmembraner under faseovergange. - M .: Nauka, 1994.
- Genis R. Biomembraner. Molekylær struktur og funktioner: oversættelse fra engelsk. = Biomembraner. Molekylær struktur og funktion (af Robert B. Genis). - 1. udgave. - M .: Mir, 1997. - ISBN 5-03-002419-0
- Ivanov V. G., Berestovsky T. N. lipid dobbeltlag af biologiske membraner. - M .: Nauka, 1982.
- Rubin A.B. Biofysik, lærebog i 2 bind. - 3. udgave, revideret og udvidet. - M .: Moscow University Press, 2004. -
Ydre cellemembran (plasmalemma, cytolemma, plasmamembran) af dyreceller dækket på ydersiden (dvs. på den side, der ikke er i kontakt med cytoplasmaet) med et lag af oligosaccharidkæder, der er kovalent bundet til membranproteiner (glykoproteiner) og i mindre grad til lipider (glykolipider). Denne kulhydratbelægning af membranen kaldes glykokalyx. Formålet med glycocalyx er endnu ikke særlig klart; der er en antagelse om, at denne struktur deltager i processerne for intercellulær genkendelse.
I planteceller oven på den ydre cellemembran er et tæt celluloselag med porer, hvorigennem kommunikationen foregår mellem naboceller gennem cytoplasmatiske broer.
Celler svampe oven på plasmalemmaet - et tæt lag kitin.
På bakterie – mureina.
Egenskaber af biologiske membraner
1. Evne til selv at samle efter destruktive påvirkninger. Denne egenskab bestemmes af de fysisk-kemiske egenskaber ved fosfolipidmolekyler, som i en vandig opløsning samles, således at de hydrofile ender af molekylerne vender udad, og de hydrofobe ender indad. Proteiner kan inkorporeres i færdiglavede fosfolipidlag. Evnen til selv at samle er afgørende på celleniveau.
2. Semi-permeabilitet(selektivitet i transmissionen af ioner og molekyler). Sikrer opretholdelsen af konstansen af den ioniske og molekylære sammensætning i cellen.
3. Membranfluiditet. Membraner er ikke stive strukturer; de svinger konstant på grund af de roterende og oscillerende bevægelser af lipid- og proteinmolekyler. Dette giver en høj hastighed af enzymatiske og andre kemiske processer i membranerne.
4. Fragmenter af membraner har ikke frie ender, da de er lukket i bobler.
Funktioner af den ydre cellemembran (plasmalemma)
Plasmalemmaets hovedfunktioner er som følger: 1) barriere, 2) receptor, 3) udveksling, 4) transport.
1. barriere funktion. Det kommer til udtryk i det faktum, at plasmalemmaet begrænser cellens indhold, adskiller det fra det ydre miljø, og intracellulære membraner deler cytoplasmaet i separate reaktionære rum.
2. receptor funktion. En af plasmalemmaets vigtigste funktioner er at sikre kommunikation (forbindelse) af cellen med det ydre miljø gennem det receptorapparat, der er til stede i membranerne, som har en protein- eller glykoproteinkarakter. Hovedfunktionen af plasmalemmaets receptorformationer er genkendelsen af eksterne signaler, på grund af hvilke cellerne er korrekt orienteret og danner væv i differentieringsprocessen. Aktiviteten af forskellige regulatoriske systemer, såvel som dannelsen af et immunrespons, er forbundet med receptorfunktionen.
udvekslingsfunktion bestemmes af indholdet af enzymproteiner i biologiske membraner, som er biologiske katalysatorer. Deres aktivitet varierer afhængigt af mediets pH, temperatur, tryk, koncentrationen af både substratet og selve enzymet. Enzymer bestemmer intensiteten af nøglereaktioner stofskifte, samt orientering.
Transportfunktion af membraner. Membranen giver selektiv penetration ind i cellen og fra cellen ind i miljøet af forskellige kemikalier. Transporten af stoffer er nødvendig for at opretholde den passende pH i cellen, den rette ionkoncentration, som sikrer effektiviteten af cellulære enzymer. Transport leverer næringsstoffer, der tjener som energikilde, samt materiale til dannelsen af forskellige cellulære komponenter. Det bestemmer fjernelsen af giftigt affald fra cellen, udskillelsen af forskellige nyttige stoffer og skabelsen af ioniske gradienter, der er nødvendige for nerve- og muskelaktivitet Ændringer i hastigheden af overførsel af stoffer kan føre til forstyrrelser i bioenergetiske processer, vand-saltmetabolisme , excitabilitet og andre processer. Korrektion af disse ændringer ligger til grund for virkningen af mange lægemidler.
Der er to hovedmåder, hvorpå stoffer kommer ind i cellen og ud af cellen i det ydre miljø;
passiv transport,
aktiv transport.
Passiv transport går langs gradienten af kemisk eller elektrokemisk koncentration uden forbrug af ATP-energi. Hvis molekylet af det transporterede stof ikke har nogen ladning, så bestemmes retningen af passiv transport kun af forskellen i koncentrationen af dette stof på begge sider af membranen (kemisk koncentrationsgradient). Hvis molekylet er ladet, så påvirkes dets transport af både den kemiske koncentrationsgradient og den elektriske gradient (membranpotentiale).
Begge gradienter udgør tilsammen en elektrokemisk gradient. Passiv transport af stoffer kan udføres på to måder: simpel diffusion og faciliteret diffusion.
Med simpel diffusion saltioner og vand kan trænge gennem de selektive kanaler. Disse kanaler er dannet af nogle transmembrane proteiner, der danner ende-til-ende transportveje, der er åbne permanent eller kun i kort tid. Gennem de selektive kanaler trænger forskellige molekyler ind, som har den størrelse og ladning, der svarer til kanalerne.
Der er en anden måde til simpel diffusion - dette er diffusionen af stoffer gennem lipid-dobbeltlaget, gennem hvilket fedtopløselige stoffer og vand let passerer. Lipid-dobbeltlaget er uigennemtrængeligt for ladede molekyler (ioner), og samtidig kan uladede små molekyler frit diffundere, og jo mindre molekylet er, jo hurtigere transporteres det. Den ret høje hastighed af vanddiffusion gennem lipid-dobbeltlaget skyldes netop den lille størrelse af dets molekyler og fraværet af en ladning.
Med lettet diffusion proteiner er involveret i transporten af stoffer - bærere, der arbejder efter princippet om "ping-pong". I dette tilfælde eksisterer proteinet i to konformationelle tilstande: i "pong"-tilstanden er bindingsstederne for det transporterede stof åbne på ydersiden af dobbeltlaget, og i "ping"-tilstanden er de samme steder åbne på den anden side. side. Denne proces er reversibel. Fra hvilken side bindingsstedet for et stof vil være åbent på et givet tidspunkt afhænger af koncentrationsgradienten af dette stof.
På denne måde passerer sukkerarter og aminosyrer gennem membranen.
Med faciliteret diffusion stiger transporthastigheden af stoffer betydeligt i sammenligning med simpel diffusion.
Ud over bærerproteiner er nogle antibiotika, såsom gramicidin og valinomycin, involveret i faciliteret diffusion.
Fordi de sørger for iontransport, kaldes de ionoforer.
Aktiv transport af stoffer i cellen. Denne type transport kommer altid med omkostningerne til energi. Den energikilde, der er nødvendig for aktiv transport, er ATP. Et karakteristisk træk ved denne type transport er, at den udføres på to måder:
ved hjælp af enzymer kaldet ATPaser;
transport i membranemballage (endocytose).
I den ydre cellemembran indeholder enzymproteiner såsom ATPaser, hvis funktion er at sørge for aktiv transport ioner mod en koncentrationsgradient. Da de sørger for transport af ioner, kaldes denne proces en ionpumpe.
Der er fire hovediontransportsystemer i dyrecellen. Tre af dem giver overførsel gennem biologiske membraner: Na + og K +, Ca +, H +, og den fjerde - overførsel af protoner under driften af den mitokondrielle respirationskæde.
Et eksempel på en aktiv iontransportmekanisme er natrium-kalium pumpe i dyreceller. Det opretholder en konstant koncentration af natrium- og kaliumioner i cellen, hvilket adskiller sig fra koncentrationen af disse stoffer i miljøet: Normalt er der færre natriumioner i cellen end i miljøet, og mere kalium.
Som et resultat, ifølge lovene om simpel diffusion, har kalium en tendens til at forlade cellen, og natrium diffunderer ind i cellen. I modsætning til simpel diffusion pumper natrium-kalium-pumpen konstant natrium ud af cellen og injicerer kalium: For tre molekyler natrium, der smides ud, er der to kaliummolekyler, der indføres i cellen.
Denne transport af natrium-kalium-ioner sikres af det ATP-afhængige enzym, som er lokaliseret i membranen på en sådan måde, at det trænger ind i hele dens tykkelse Natrium og ATP kommer ind i dette enzym fra indersiden af membranen, og kalium fra membranen. uden for.
Overførslen af natrium og kalium over membranen sker som følge af konformationsændringer, som den natrium-kalium-afhængige ATPase gennemgår, som aktiveres, når koncentrationen af natrium inde i cellen eller kalium i miljøet stiger.
ATP-hydrolyse er påkrævet for at drive denne pumpe. Denne proces leveres af det samme enzym natrium-kalium-afhængige ATP-ase. Samtidig bruges mere end en tredjedel af den ATP, der forbruges af en dyrecelle i hvile, på driften af natrium-kalium-pumpen.
Overtrædelse af den korrekte funktion af natrium-kaliumpumpen fører til forskellige alvorlige sygdomme.
Effektiviteten af denne pumpe overstiger 50%, hvilket ikke opnås af de mest avancerede maskiner skabt af mennesker.
Mange aktive transportsystemer drives af energi lagret i ioniske gradienter snarere end af direkte hydrolyse af ATP. Alle fungerer som cotransportsystemer (der letter transporten af forbindelser med lav molekylvægt). For eksempel bestemmes den aktive transport af visse sukkerarter og aminosyrer ind i dyreceller af natriumiongradienten, og jo højere natriumiongradienten er, desto større er glucoseabsorptionshastigheden. Omvendt, hvis koncentrationen af natrium i det intercellulære rum falder markant, stopper glukosetransporten. I dette tilfælde skal natrium slutte sig til det natriumafhængige glukosebærerprotein, som har to bindingssteder: det ene for glucose, det andet for natrium. Natriumioner, der trænger ind i cellen, bidrager til introduktionen af bærerproteinet i cellen sammen med glucose. Natriumioner, der er kommet ind i cellen sammen med glukose, pumpes ud igen af den natrium-kalium-afhængige ATPase, som ved at opretholde natriumkoncentrationsgradienten indirekte styrer glukosetransporten.
Transport af stoffer i membranemballage. Store molekyler af biopolymerer kan praktisk talt ikke trænge gennem plasmalemmaet af nogen af de ovenfor beskrevne mekanismer til transport af stoffer ind i cellen. De fanges af cellen og absorberes i membranpakken, som kaldes endocytose. Sidstnævnte er formelt opdelt i fagocytose og pinocytose. Indfangningen af faste partikler af cellen er fagocytose og flydende - pinocytose. Under endocytose observeres følgende stadier:
modtagelse af det absorberede stof på grund af receptorer i cellemembranen;
invagination af membranen med dannelsen af en boble (vesikler);
adskillelse af den endocytiske vesikel fra membranen med energiforbrug - fagosomdannelse og restaurering af membranintegritet;
Fusion af fagosom med lysosom og dannelse fagolysosomer (fordøjelsesvakuole) hvor fordøjelsen af absorberede partikler finder sted;
fjernelse af ufordøjet materiale i fagolysosomet fra cellen ( eksocytose).
I dyreriget endocytose er en karakteristisk måde at fodre mange encellede organismer på (for eksempel i amøber), og blandt flercellede organismer findes denne type fordøjelse af fødepartikler i endodermale celler i coelenterater. Hvad angår pattedyr og mennesker, har de et reticulo-histio-endotelsystem af celler med evnen til endocytose. Eksempler er blodleukocytter og lever Kupffer-celler. Sidstnævnte beklæder de såkaldte sinusformede kapillærer i leveren og fanger forskellige fremmede partikler suspenderet i blodet. Exocytose- dette er også en måde at fjerne det substrat, der udskilles af den, fra cellen i en flercellet organisme, hvilket er nødvendigt for funktionen af andre celler, væv og organer.

