30.09.2019
Mitä solukalvon reikiä kutsutaan? Kalvot: niiden rakenne ja toiminta
Kaikki elävät organismit on solun rakenteesta riippuen jaettu kolmeen ryhmään (katso kuva 1):
1. Prokaryootit (ei-ydin)
2. Eukaryootit (ydin)
3. Virukset (ei-soluiset)
Riisi. 1. Elävät organismit
Tällä oppitunnilla alamme tutkia eukaryoottisten organismien solujen rakennetta, joihin kuuluvat kasvit, sienet ja eläimet. Niiden solut ovat suurimpia ja monimutkaisimpia verrattuna prokaryoottisoluihin.
Kuten tiedät, solut pystyvät itsenäiseen toimintaan. Ne voivat vaihtaa ainetta ja energiaa ympäristön kanssa sekä kasvaa ja lisääntyä, joten solun sisäinen rakenne on hyvin monimutkainen ja riippuu ensisijaisesti tehtävästä, jota solu suorittaa monisoluisessa organismissa.
Kaikkien solujen rakennusperiaatteet ovat samat. Jokaisessa eukaryoottisolussa voidaan erottaa seuraavat pääosat (katso kuva 2):
1. Ulkokalvo, joka erottaa solun sisällön ulkoisesta ympäristöstä.
2. Sytoplasma, jossa on organelleja.

Riisi. 2. Eukaryoottisolun pääosat
Termiä "kalvo" ehdotettiin noin sata vuotta sitten kuvaamaan solun rajoja, mutta elektronimikroskopian kehittyessä kävi selväksi, että solukalvo on osa solun rakenneosia.
Vuonna 1959 J. D. Robertson muotoili elementaarisen kalvohypoteesin, jonka mukaan eläinten ja kasvien solukalvot rakennetaan saman tyypin mukaan.
Vuonna 1972 Singer ja Nicholson ehdottivat sitä, mikä on tällä hetkellä yleisesti hyväksyttyä. Tämän mallin mukaan minkä tahansa kalvon perusta on kaksinkertainen fosfolipidikerros.
Fosfolipideissä (yhdisteet, jotka sisältävät fosfaattiryhmän) molekyylit koostuvat polaarisesta päästä ja kahdesta ei-polaarisesta hännästä (katso kuva 3).

Riisi. 3. Fosfolipidi
Fosfolipidikaksoiskerroksessa hydrofobiset rasvahappojäännökset osoittavat sisäänpäin, kun taas hydrofiiliset päät, mukaan lukien fosforihappojäännös, ovat ulospäin (katso kuva 4).

Riisi. 4. Fosfolipidikaksoiskerros
Fosfolipidikaksoiskerros esitetään dynaamisena rakenteena, lipidit voivat liikkua vaihtaen sijaintiaan.
Kaksinkertainen lipidien kerros huolehtii kalvon sulkutoiminnosta, joka estää solun sisällön leviämisen ja estää myrkyllisten aineiden pääsyn soluun.
Rajakalvon läsnäolo solun ja ympäristön välillä tiedettiin kauan ennen elektronimikroskoopin tuloa. Fysikaaliset kemistit kielsivät plasmakalvon olemassaolon ja uskoivat, että elävän kolloidisen sisällön ja ympäristön välillä oli rajapinta, mutta Pfeffer (saksalainen kasvitieteilijä ja kasvifysiologi) vahvisti sen olemassaolon vuonna 1890.
Viime vuosisadan alussa Overton (britlantilainen fysiologi ja biologi) havaitsi, että monien aineiden tunkeutumisnopeus punasoluihin on suoraan verrannollinen niiden rasvaliukoisuuteen. Tältä osin tiedemies ehdotti, että kalvo sisältää suuren määrän lipidejä ja aineita, jotka liukenevat siihen, kulkevat sen läpi ja joutuvat kalvon toiselle puolelle.
Vuonna 1925 Gorter ja Grendel (amerikkalaiset biologit) eristivät lipidejä punasolujen solukalvosta. Tuloksena saadut lipidit jakautuivat veden pinnalle yhden molekyylin paksuudella. Kävi ilmi, että lipidikerroksen miehittämä pinta-ala on kaksi kertaa suurempi kuin itse punasolun pinta-ala. Siksi nämä tutkijat päättelivät, että solukalvo ei koostu yhdestä, vaan kahdesta lipidikerroksesta.
Dawson ja Danielli (englantilaiset biologit) ehdottivat vuonna 1935, että solukalvoissa bimolekulaarinen lipidikerros on suljettu kahden proteiinimolekyylikerroksen väliin (katso kuva 5).

Riisi. 5. Dawsonin ja Daniellin ehdottama kalvomalli
Elektronimikroskoopin tultua mahdolliseksi tutustua kalvon rakenteeseen, ja sitten havaittiin, että eläin- ja kasvisolujen kalvot näyttävät kolmikerroksiselta rakenteelta (ks. kuva 6).

Riisi. 6. Solukalvo mikroskoopin alla
Vuonna 1959 biologi J. D. Robertson, yhdistäen tuolloin saatavilla olevat tiedot, esitti hypoteesin "alkukalvon" rakenteesta, jossa hän oletti rakenteen, joka on yhteinen kaikille biologisille kalvoille.
Robertsonin postulaatit "alkukalvon" rakenteesta
1. Kaikki kalvot ovat noin 7,5 nm paksuja.
2. Elektronimikroskoopissa ne kaikki näyttävät olevan kolmikerroksisia.
3. Kolmikerroksinen näkymä kalvosta on seurausta täsmälleen proteiinien ja polaaristen lipidien järjestelystä, joka saatiin aikaan Dawsonin ja Daniellin mallissa - keskuslipidikaksoiskerros on suljettu kahden proteiinikerroksen väliin.
Tämä hypoteesi "alkukalvon" rakenteesta on kokenut useita muutoksia, ja vuonna 1972 sen esitti kalvon nestemosaiikkimalli(katso kuva 7), joka on nyt yleisesti hyväksytty.

Riisi. 7. Kalvon nestemosaiikkimalli
Proteiinimolekyylit upotetaan kalvon lipidikaksoiskerrokseen, ne muodostavat liikkuvan mosaiikin. Sen mukaan, miten ne ovat vuorovaikutuksessa lipidikaksoiskerroksen kanssa, proteiinit voidaan jakaa:
- pinnallinen (tai perifeerinen) kalvoproteiinit, jotka liittyvät lipidikaksoiskerroksen hydrofiiliseen pintaan;
- integraali (kalvo) kaksikerroksen hydrofobiselle alueelle upotettuja proteiineja.
Integraalit proteiinit eroavat toisistaan siinä, missä määrin ne uppoavat kaksoiskerroksen hydrofobiseen alueeseen. Ne voidaan upottaa kokonaan kiinteä) tai osittain veden alla ( puolikiinteä) ja voi myös tunkeutua kalvon läpi ( kalvon läpäisevä).
Kalvoproteiinit voidaan jakaa kahteen ryhmään niiden toimintojen mukaan:
- rakenteellinen proteiinit. Ne ovat osa solukalvoja ja osallistuvat niiden rakenteen ylläpitämiseen.
- dynaaminen proteiinit. Ne sijaitsevat kalvoilla ja osallistuvat niillä tapahtuviin prosesseihin.
Dynaamisia proteiineja on kolme luokkaa.
1. Reseptori. Näiden proteiinien avulla solu havaitsee pinnallaan erilaisia vaikutuksia. Toisin sanoen ne sitovat spesifisesti yhdisteitä, kuten hormoneja, välittäjäaineita, myrkkyjä kalvon ulkopuolella, mikä toimii signaalina muuttaa erilaisia prosesseja solun sisällä tai itse kalvossa.
2. Kuljetus. Nämä proteiinit kuljettavat tiettyjä aineita kalvon läpi, ne muodostavat myös kanavia, joiden kautta erilaisia ioneja kuljetetaan soluun ja sieltä pois.
3. Entsymaattinen. Nämä ovat entsyymiproteiineja, jotka sijaitsevat kalvossa ja ovat mukana erilaisissa kemiallisissa prosesseissa.
Aineiden kulkeutuminen kalvon läpi
Lipidikaksoiskerrokset ovat suurelta osin läpäisemättömiä monille aineille, joten aineiden kuljettamiseen kalvon läpi tarvitaan suuri määrä energiaa ja myös erilaisten rakenteiden muodostumista.
Kuljetuksia on kahta tyyppiä: passiivinen ja aktiivinen.
Passiivinen kuljetus
Passiivinen kuljetus on molekyylien liikettä pitoisuusgradienttia pitkin. Toisin sanoen se määräytyy vain siirretyn aineen pitoisuuden eron perusteella kalvon vastakkaisilla puolilla ja se suoritetaan ilman energiankulutusta.
Passiivista kuljetusta on kahta tyyppiä:
- yksinkertainen diffuusio(katso kuvio 8), joka tapahtuu ilman kalvoproteiinin osallistumista. Yksinkertaisen diffuusion mekanismi on kaasujen (happi ja hiilidioksidi), veden ja joidenkin yksinkertaisten orgaanisten ionien siirto kalvon läpi. Yksinkertainen diffuusio on hidasta.

Riisi. 8. Yksinkertainen diffuusio
- helpotettu diffuusio(katso kuvio 9) eroaa yksinkertaisesta siinä, että se tapahtuu kantajaproteiinien osallistuessa. Tämä prosessi on spesifinen ja etenee nopeammin kuin yksinkertainen diffuusio.

Riisi. 9. Helpotettu leviäminen
Tunnetaan kahdenlaisia kalvonkuljetusproteiineja: kantajaproteiinit (translokaasit) ja kanavaa muodostavat proteiinit. Kuljetusproteiinit sitovat tiettyjä aineita ja kuljettavat niitä kalvon läpi pitoisuusgradienttiaan pitkin, ja näin ollen tämä prosessi, kuten yksinkertaisessa diffuusiossa, ei vaadi ATP-energian kuluttamista.
Ruokahiukkaset eivät pääse kalvon läpi, vaan ne tulevat soluun endosytoosin kautta (ks. kuva 10). Endosytoosin aikana plasmakalvo muodostaa invaginaatioita ja kasvaimia, vangitsee kiinteän ruokapartikkelin. Ruokaboluksen ympärille muodostuu tyhjiö (tai vesikkeli), joka sitten irtoaa plasmakalvosta ja tyhjiön kiinteä partikkeli on solun sisällä.

Riisi. 10. Endosytoosi
Endosytoosia on kahta tyyppiä.
1. Fagosytoosi- kiinteiden hiukkasten imeytyminen. Fagosytoosia suorittavia erikoistuneita soluja kutsutaan fagosyytit.
2. pinosytoosi- nestemäisen materiaalin (liuos, kolloidinen liuos, suspensiot) imeytyminen.
Eksosytoosi(katso kuvio 11) - endosytoosiin päinvastainen prosessi. Solussa syntetisoituvat aineet, kuten hormonit, pakataan solukalvoon sopiviin kalvorakkuloihin, upotetaan siihen ja rakkulan sisältö poistetaan solusta. Samalla tavalla solu pääsee eroon tarpeettomista aineenvaihduntatuotteista.

Riisi. 11. Eksosytoosi
aktiivinen kuljetus
Toisin kuin helpotettu diffuusio, aktiivinen kuljetus on aineiden liikkumista pitoisuusgradienttia vasten. Tässä tapauksessa aineet siirtyvät alueelta, jolla on pienempi pitoisuus, alueelle, jolla on suurempi pitoisuus. Koska tällainen liike tapahtuu päinvastaiseen suuntaan kuin normaali diffuusio, kennon on kulutettava energiaa tässä prosessissa.
Esimerkeistä aktiivisesta kuljetuksesta on parhaiten tutkittu ns. natrium-kaliumpumppu. Tämä pumppu pumppaa natriumioneja ulos solusta ja pumppaa kaliumioneja soluun ATP:n energialla.
1. Rakenteellinen (solukalvo erottaa solun ympäristöstä).
2. Kuljetus (aineet kuljetetaan solukalvon läpi, ja solukalvo on erittäin selektiivinen suodatin).
3. Reseptori (kalvon pinnalla sijaitsevat reseptorit havaitsevat ulkoiset vaikutukset, välittävät tämän tiedon soluun, jolloin se pystyy reagoimaan nopeasti ympäristön muutoksiin).
Edellä lueteltujen lisäksi kalvo suorittaa myös metabolista ja energiaa muuntavaa toimintaa.
metabolinen toiminta
Biologiset kalvot osallistuvat suoraan tai epäsuorasti aineiden aineenvaihduntaprosesseihin solussa, koska useimmat entsyymit liittyvät kalvoihin.
Entsyymien lipidiympäristö kalvossa luo tietyt edellytykset niiden toiminnalle, asettaa rajoituksia kalvoproteiinien aktiivisuudelle ja siten säätelee aineenvaihduntaprosesseja.
Energian muunnostoiminto
Monien biokalvojen tärkein tehtävä on yhden energiamuodon muuntaminen toiseksi.
Energiaa muuntavia kalvoja ovat mitokondrioiden sisäiset kalvot, kloroplastien tylakoidit (katso kuva 12).

Riisi. 12. Mitokondriot ja kloroplastit
Bibliografia
- Kamensky A.A., Kriksunov E.A., Pasechnik V.V. Yleinen biologia 10-11 luokka Bustard, 2005.
- Biologia. Luokka 10. Yleinen biologia. Perustaso / P.V. Izhevsky, O.A. Kornilova, T.E. Loshchilin ja muut - 2. painos, tarkistettu. - Ventana-Graf, 2010. - 224 sivua.
- Belyaev D.K. Biologia 10-11 luokka. Yleinen biologia. Perustaso. - 11. painos, stereotypia. - M.: Koulutus, 2012. - 304 s.
- Agafonova I.B., Zakharova E.T., Sivoglazov V.I. Biologia 10-11 luokka. Yleinen biologia. Perustaso. - 6. painos, lisäys. - Bustard, 2010. - 384 s.
- Ayzdorov.ru ().
- Youtube.com().
- Doctor-v.ru ().
- Animals-world.ru ().
Kotitehtävät
- Mikä on solukalvon rakenne?
- Mitkä ovat lipidien ominaisuudet muodostaa kalvoja?
- Minkä toimintojen ansiosta proteiinit voivat osallistua aineiden kuljettamiseen kalvon läpi?
- Luettele plasmakalvon tehtävät.
- Kuinka passiivinen kuljetus tapahtuu kalvon läpi?
- Kuinka aktiivinen kuljetus tapahtuu kalvon läpi?
- Mikä on natrium-kaliumpumpun tehtävä?
- Mikä on fagosytoosi, pinosytoosi?
Organismien, samoin kuin kasvien, eläinten ja ihmisten rakenteen tutkimus on biologian haara, jota kutsutaan sytologiaksi. Tutkijat ovat havainneet, että sen sisällä olevan solun sisältö on melko monimutkaista. Sitä ympäröi ns. pintalaitteisto, joka sisältää ulomman solukalvon, suprakalvorakenteet: glykokaliksi ja mikrofilamentit, pelikulit ja mikrotubulukset, jotka muodostavat sen submembraanikompleksin.
Tässä artikkelissa tutkimme ulomman solukalvon rakennetta ja toimintoja, jotka ovat osa erityyppisten solujen pintalaitteistoa.
Mitkä ovat ulomman solukalvon tehtävät?
Kuten aiemmin on kuvattu, ulkokalvo on osa kunkin solun pintalaitteistoa, joka erottaa onnistuneesti sen sisäisen sisällön ja suojaa soluorganelleja haitallisilta ympäristöolosuhteilta. Toinen tehtävä on varmistaa aineiden vaihto solun sisällön ja kudosnesteen välillä, joten ulompi solukalvo kuljettaa molekyylejä ja ioneja, jotka tulevat sytoplasmaan, ja auttaa myös poistamaan myrkkyjä ja ylimääräisiä myrkyllisiä aineita solusta.

Solukalvon rakenne
Erityyppisten solujen kalvot tai plasmalemmat ovat hyvin erilaisia toisistaan. Pääasiassa kemiallinen rakenne sekä lipidien, glykoproteiinien, proteiinien suhteellinen pitoisuus niissä ja vastaavasti niissä olevien reseptorien luonne. Ulkoinen, joka määräytyy ensisijaisesti glykoproteiinien yksilöllisen koostumuksen perusteella, osallistuu ympäristön ärsykkeiden tunnistamiseen ja itse solun reaktioihin niiden toimintaan. Jotkut virukset voivat olla vuorovaikutuksessa solukalvojen proteiinien ja glykolipidien kanssa, minkä seurauksena ne tunkeutuvat soluun. Herpes- ja influenssavirukset voivat rakentaa suojakuorensa.
Ja virukset ja bakteerit, niin sanotut bakteriofagit, kiinnittyvät solukalvoon ja liuottavat sen kosketuspisteessä erityisen entsyymin avulla. Sitten viruksen DNA-molekyyli siirtyy muodostuneeseen reikään.
Eukaryoottien plasmakalvon rakenteen piirteet
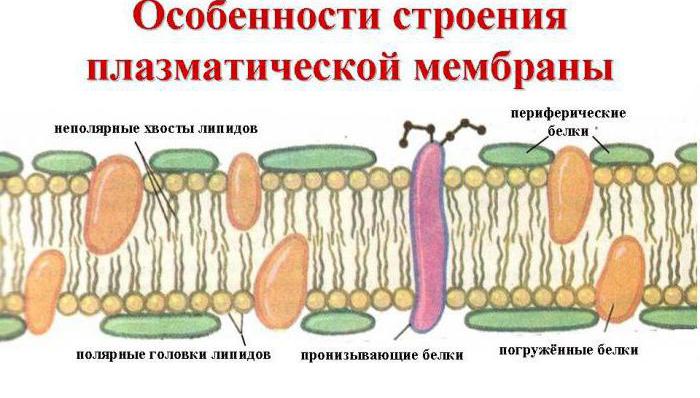
Muista, että ulompi solukalvo suorittaa kuljetustoimintoa, eli aineiden siirtoa siihen ja sieltä ulos ulkoiseen ympäristöön. Tällaisen prosessin suorittamiseksi tarvitaan erityinen rakenne. Itse asiassa plasmalemma on vakio, universaali pintalaitteiston järjestelmä kaikille. Tämä on ohut (2-10 Nm), mutta melko tiheä monikerroksinen kalvo, joka peittää koko kennon. Sen rakennetta tutkivat vuonna 1972 sellaiset tiedemiehet kuin D. Singer ja G. Nicholson, ja he loivat myös nestemosaiikkimallin solukalvosta.
Tärkeimmät sen muodostavat kemialliset yhdisteet ovat proteiinien ja tiettyjen fosfolipidien järjestettyjä molekyylejä, jotka ovat kietoutuneet nestemäiseen lipidiympäristöön ja muistuttavat mosaiikkia. Siten solukalvo koostuu kahdesta kerroksesta lipidejä, joiden ei-polaariset hydrofobiset "hännät" sijaitsevat kalvon sisällä ja polaariset hydrofiiliset päät ovat solun sytoplasmaan ja solujen väliseen nesteeseen päin.
Lipidikerroksen läpäisevät suuret proteiinimolekyylit, jotka muodostavat hydrofiilisiä huokosia. Niiden kautta kuljetetaan glukoosin ja mineraalisuolojen vesiliuoksia. Jotkut proteiinimolekyylit sijaitsevat sekä plasmalemman ulko- että sisäpinnalla. Siten kaikkien ytimiä sisältävien organismien solujen ulkoisella solukalvolla on hiilihydraattimolekyylejä, jotka on sitoutunut kovalenttisilla sidoksilla glykolipidien ja glykoproteiinien kanssa. Hiilihydraattien pitoisuus solukalvoissa vaihtelee välillä 2-10%.

Prokaryoottisten organismien plasmalemman rakenne
Prokaryoottien ulompi solukalvo suorittaa samanlaisia tehtäviä kuin ydinorganismien solujen plasmakalvot, nimittäin: havaitsee ja välittää ulkoisesta ympäristöstä tulevaa tietoa, kuljettaa ioneja ja liuoksia soluun ja sieltä pois sekä suojaa soluja. sytoplasma ulkopuolelta tulevilta vierailta reagensseilta. Se voi muodostaa mesosomeja - rakenteita, jotka syntyvät, kun plasmalemma työntyy soluun. Ne voivat sisältää entsyymejä, jotka osallistuvat prokaryoottien metabolisiin reaktioihin, esimerkiksi DNA:n replikaatioon, proteiinisynteesiin.
Mesosomit sisältävät myös redox-entsyymejä, kun taas fotosynteettiset aineet sisältävät bakterioklorofylliä (bakteerissa) ja fikobiliinia (syanobakteereissa).
Ulkokalvojen rooli solujen välisissä kontakteissa
Jatkamalla vastausta kysymykseen, mitä toimintoja ulompi solukalvo suorittaa, katsotaanpa sen roolia kasvisoluissa: Kasvisoluissa ulomman solukalvon seinämiin muodostuu huokoset, jotka siirtyvät selluloosakerrokseen. Niiden kautta solun sytoplasman poistuminen ulkopuolelle on mahdollista, tällaisia ohuita kanavia kutsutaan plasmodesmaiksi.

Niiden ansiosta yhteys vierekkäisten kasvisolujen välillä on erittäin vahva. Ihmis- ja eläinsoluissa vierekkäisten solukalvojen välisiä kosketuskohtia kutsutaan desmosomeiksi. Ne ovat ominaisia endoteelisoluille ja epiteelisoluille, ja niitä löytyy myös sydänlihassoluista.
Plasmalemman apumuodostelmat
Ymmärtääkseen, miten kasvisolut eroavat eläimistä, se auttaa tutkimaan niiden plasmakalvojen rakenteellisia piirteitä, jotka riippuvat siitä, mitä toimintoja ulompi solukalvo suorittaa. Sen yläpuolella eläinsoluissa on glykokaliksikerros. Sen muodostavat polysakkaridimolekyylit, jotka liittyvät ulkoisen solukalvon proteiineihin ja lipideihin. Glykokalyyksin ansiosta solujen välillä tapahtuu adheesiota (tarttumista), mikä johtaa kudosten muodostumiseen, joten se osallistuu plasmalemman signalointitoimintoon - ympäristön ärsykkeiden tunnistamiseen.
Miten tiettyjen aineiden passiivinen kuljetus tapahtuu solukalvojen läpi
Kuten aiemmin mainittiin, ulompi solukalvo on mukana prosessissa kuljettaa aineita solun ja ulkoisen ympäristön välillä. Plasmalemman kautta kulkee kahta tyyppiä: passiivinen (diffuusio) ja aktiivinen kuljetus. Ensimmäinen sisältää diffuusion, helpotetun diffuusion ja osmoosin. Aineiden liikkuminen pitoisuusgradienttia pitkin riippuu ensisijaisesti solukalvon läpi kulkevien molekyylien massasta ja koosta. Esimerkiksi pienet ei-polaariset molekyylit liukenevat helposti plasmalemman keskilipidikerrokseen, liikkuvat sen läpi ja päätyvät sytoplasmaan.

Suuret orgaanisten aineiden molekyylit tunkeutuvat sytoplasmaan erityisten kantajaproteiinien avulla. Ne ovat lajispesifisiä ja yhdessä hiukkasen tai ionin kanssa kuljettavat ne passiivisesti kalvon läpi pitoisuusgradienttia pitkin (passiivinen kuljetus) kuluttamatta energiaa. Tämä prosessi on sellaisen plasmalemman ominaisuuden taustalla kuin selektiivinen läpäisevyys. Prosessissa ATP-molekyylien energiaa ei käytetä, vaan solu säästää sen muihin aineenvaihduntareaktioihin.
Kemiallisten yhdisteiden aktiivinen kuljetus plasmalemman poikki
Koska ulompi solukalvo varmistaa molekyylien ja ionien siirtymisen ulkoisesta ympäristöstä soluun ja takaisin, on mahdollista poistaa dissimilaatiotuotteet, jotka ovat toksiineja, ulospäin, eli solujen väliseen nesteeseen. esiintyy pitoisuusgradienttia vastaan ja vaatii energian käyttöä ATP-molekyylien muodossa. Se sisältää myös kantajaproteiineja, joita kutsutaan ATPaaseiksi, jotka ovat myös entsyymejä.

Esimerkki tällaisesta kuljetuksesta on natrium-kaliumpumppu (natrium-ionit siirtyvät sytoplasmasta ulkoiseen ympäristöön ja kalium-ionit pumpataan sytoplasmaan). Suolen ja munuaisten epiteelisolut pystyvät siihen. Tämän siirtomenetelmän lajikkeet ovat pinosytoosi- ja fagosytoosiprosessit. Siten tutkittuaan, mitä toimintoja ulompi solukalvo suorittaa, voidaan todeta, että heterotrofiset protistit sekä korkeampien eläinorganismien solut, esimerkiksi leukosyytit, kykenevät pino- ja fagosytoosiin.
Biosähköiset prosessit solukalvoissa
On todettu, että plasmalemman ulkopinnan (se on positiivisesti varautunut) ja negatiivisesti varautuneen sytoplasman parietaalikerroksen välillä on potentiaaliero. Sitä kutsuttiin lepopotentiaaliksi, ja se on luontainen kaikille eläville soluille. Ja hermokudoksella ei ole vain lepopotentiaalia, vaan se pystyy myös johtamaan heikkoja biovirtoja, jota kutsutaan viritysprosessiksi. Hermosolujen-neuronien ulkokalvot, jotka saavat ärsytystä reseptoreista, alkavat muuttaa varauksia: natriumionit tulevat massiivisesti soluun ja plasmalemman pinta muuttuu elektronegatiiviseksi. Ja sytoplasman parietaalinen kerros saa positiivisen varauksen kationien ylimäärän vuoksi. Tämä selittää, miksi hermosolun ulompi solukalvo latautuu, mikä aiheuttaa viritysprosessin taustalla olevien hermoimpulssien johtumisen.
solukalvo
Kuva solukalvosta. Pienet siniset ja valkoiset pallot vastaavat fosfolipidien hydrofobisia "päitä" ja niihin kiinnitetyt linjat vastaavat hydrofiilisiä "häntiä". Kuvassa näkyvät vain integraaliset kalvoproteiinit (punaiset pallot ja keltaiset heliksit). Keltaiset soikeat täplät kalvon sisällä - kolesterolimolekyylit Keltaisenvihreät helmiketjut kalvon ulkopuolella - oligosakkaridiketjut, jotka muodostavat glykokalyksin
Biologiseen kalvoon kuuluu myös erilaisia proteiineja: integraali (tunkeutuu kalvon läpi), puoliintegraali (toisesta päästä upotettu ulompaan tai sisempään lipidikerrokseen), pinta (sijaitsee kalvon ulkopinnalla tai sisäsivujen vieressä). Jotkut proteiinit ovat solukalvon kosketuspisteitä solun sisällä olevan sytoskeleton kanssa ja soluseinän (jos sellainen on) ulkopuolella. Jotkut integroiduista proteiineista toimivat ionikanavina, erilaisina kuljettajina ja reseptoreina.
Toiminnot
- este - tarjoaa säädellyn, selektiivisen, passiivisen ja aktiivisen aineenvaihdunnan ympäristön kanssa. Esimerkiksi peroksisomikalvo suojaa sytoplasmaa solulle vaarallisilta peroksideilta. Selektiivinen läpäisevyys tarkoittaa, että kalvon läpäisevyys eri atomeille tai molekyyleille riippuu niiden koosta, sähkövarauksesta ja kemiallisista ominaisuuksista. Selektiivinen läpäisevyys varmistaa solun ja soluosaston erottamisen ympäristöstä ja toimittaa niille tarvittavat aineet.
- kuljetus - kalvon läpi tapahtuu aineiden kuljetus soluun ja solusta ulos. Kuljetus kalvojen läpi mahdollistaa: ravinteiden toimittamisen, aineenvaihdunnan lopputuotteiden poistumisen, erilaisten aineiden erittymisen, ionigradienttien muodostumisen, optimaalisen ionien pitoisuuden ylläpidon solussa, jotka ovat välttämättömiä solujen toiminnan kannalta. soluentsyymit.
Hiukkaset, jotka eivät jostain syystä pysty läpäisemään fosfolipidikaksoiskerrosta (esimerkiksi hydrofiilisten ominaisuuksien vuoksi, koska kalvo on sisällä hydrofobinen eikä päästä hydrofiilisiä aineita läpi, tai suuren koonsa vuoksi), mutta solulle välttämättömiä , voivat tunkeutua kalvon läpi erityisten kantajaproteiinien (kuljettajien) ja kanavaproteiinien kautta tai endosytoosin kautta.
Passiivisessa kuljetuksessa aineet ylittävät lipidikaksoiskerroksen ilman energiankulutusta pitoisuusgradienttia pitkin diffuusion kautta. Tämän mekanismin muunnelma on helpotettu diffuusio, jossa tietty molekyyli auttaa ainetta kulkemaan kalvon läpi. Tällä molekyylillä voi olla kanava, jonka läpi vain yhden tyyppinen aine pääsee kulkemaan.
Aktiivinen kuljetus vaatii energiaa, koska se tapahtuu pitoisuusgradienttia vastaan. Kalvolla on erityisiä pumppuproteiineja, mukaan lukien ATPaasi, joka pumppaa aktiivisesti kaliumioneja (K +) soluun ja pumppaa natriumioneja (Na +) ulos siitä. - matriisi - tarjoaa kalvoproteiinien tietyn suhteellisen sijainnin ja suunnan, niiden optimaalisen vuorovaikutuksen.
- mekaaninen - varmistaa solun autonomian, sen solunsisäiset rakenteet sekä yhteyden muihin soluihin (kudoksissa). Soluseinillä on tärkeä rooli mekaanisen toiminnan tarjoamisessa, ja eläimillä - solujen välisellä aineella.
- energia - fotosynteesin aikana kloroplasteissa ja soluhengityksen aikana mitokondrioissa niiden kalvoissa toimivat energiansiirtojärjestelmät, joihin myös proteiinit osallistuvat;
- reseptori - jotkut kalvossa sijaitsevat proteiinit ovat reseptoreita (molekyylejä, joiden avulla solu havaitsee tiettyjä signaaleja).
Esimerkiksi veressä kiertävät hormonit vaikuttavat vain kohdesoluihin, joissa on näitä hormoneja vastaavat reseptorit. Välittäjäaineet (kemikaalit, jotka johtavat hermoimpulsseja) sitoutuvat myös kohdesolujen spesifisiin reseptoriproteiineihin. - entsymaattinen - kalvoproteiinit ovat usein entsyymejä. Esimerkiksi suoliston epiteelisolujen plasmakalvot sisältävät ruoansulatusentsyymejä.
- biopotentiaalien synnyttämisen ja johtamisen toteuttaminen.
Kalvon avulla ylläpidetään ionien vakiopitoisuutta solussa: K + -ionin pitoisuus solun sisällä on paljon korkeampi kuin sen ulkopuolella ja Na + -pitoisuus on paljon pienempi, mikä on erittäin tärkeää, koska tämä säilyttää potentiaalieron kalvon poikki ja synnyttää hermoimpulssin. - solumerkintä - kalvolla on antigeenejä, jotka toimivat markkereina - "leimat", jotka mahdollistavat solun tunnistamisen. Nämä ovat glykoproteiineja (eli proteiineja, joihin on kiinnitetty haarautuneita oligosakkaridisivuketjuja), joilla on "antennien" rooli. Lukuisten sivuketjukonfiguraatioiden ansiosta on mahdollista tehdä erityinen markkeri jokaiselle solutyypille. Markkerien avulla solut voivat tunnistaa muita soluja ja toimia niiden kanssa yhdessä esimerkiksi elimiä ja kudoksia muodostaessaan. Sen avulla immuunijärjestelmä pystyy myös tunnistamaan vieraita antigeenejä.
Biokalvojen rakenne ja koostumus
Kalvot koostuvat kolmesta lipidien luokasta: fosfolipideistä, glykolipideistä ja kolesterolista. Fosfolipidit ja glykolipidit (lipidit, joihin on kiinnittynyt hiilihydraatteja) koostuvat kahdesta pitkästä hydrofobisesta hiilivety "hännästä", jotka liittyvät varautuneeseen hydrofiiliseen "päähän". Kolesteroli jäykistää kalvoa viemällä vapaan tilan hydrofobisten lipidipyrstöjen väliin ja estämällä niitä taipumasta. Siksi kalvot, joissa on alhainen kolesterolipitoisuus, ovat joustavampia, kun taas korkean kolesterolipitoisuuden omaavat kalvot ovat jäykempiä ja hauraampia. Kolesteroli toimii myös "sulkuna", joka estää polaaristen molekyylien liikkumisen solusta ja soluun. Tärkeä osa kalvoa koostuu proteiineista, jotka läpäisevät sen ja vastaavat kalvojen erilaisista ominaisuuksista. Niiden koostumus ja suuntaus eri kalvoissa vaihtelee.
Solukalvot ovat usein epäsymmetrisiä, eli kerrokset eroavat lipidikoostumuksesta, yksittäisen molekyylin siirtymisestä kerroksesta toiseen (ns. varvastossu) on vaikea.
Kalvoorganellit
Nämä ovat suljettuja yksittäisiä tai toisiinsa liittyviä sytoplasman osia, jotka on erotettu hyaloplasmasta kalvoilla. Yksikalvoisia organelleja ovat endoplasminen verkkokalvo, Golgin laite, lysosomit, vakuolit, peroksisomit; kaksikalvoiseen - ydin, mitokondriot, plastidit. Erilaisten organellien kalvojen rakenne eroaa lipidien ja kalvoproteiinien koostumuksesta.
Valikoiva läpäisevyys
Solukalvoilla on selektiivinen läpäisevyys: glukoosi, aminohapot, rasvahapot, glyseroli ja ionit diffundoituvat hitaasti niiden läpi, ja kalvot itse säätelevät tätä prosessia aktiivisesti jossain määrin - jotkut aineet kulkevat läpi, kun taas toiset eivät. Aineiden pääsylle soluun tai niiden poistamiselle solusta ulos on neljä päämekanismia: diffuusio, osmoosi, aktiivinen kuljetus ja ekso- tai endosytoosi. Kaksi ensimmäistä prosessia ovat luonteeltaan passiivisia, eli ne eivät vaadi energiaa; kaksi viimeistä ovat aktiivisia energiankulutukseen liittyviä prosesseja.
Kalvon selektiivinen läpäisevyys passiivisen kuljetuksen aikana johtuu erityisistä kanavista - integraalisista proteiineista. Ne tunkeutuvat kalvon läpi ja läpi muodostaen eräänlaisen käytävän. Alkuaineilla K, Na ja Cl on omat kanavansa. Pitoisuusgradientin suhteen näiden alkuaineiden molekyylit liikkuvat soluun sisään ja ulos. Ärsyttyessä natriumionikanavat avautuvat ja natriumioneja virtaa jyrkästi soluun. Tämä johtaa epätasapainoon kalvopotentiaalissa. Sen jälkeen kalvopotentiaali palautuu. Kaliumkanavat ovat aina auki, niiden kautta kaliumionit tulevat hitaasti soluun.
Katso myös
Kirjallisuus
- Antonov V. F., Smirnova E. N., Shevchenko E. V. Lipidikalvot faasimuutosten aikana. - M .: Nauka, 1994.
- Gennis R. Biokalvot. Molekyylirakenne ja toiminnot: käännös englannista. = Biokalvot. Molekyylirakenne ja toiminta (Robert B. Gennis). - 1. painos. - M .: Mir, 1997. - ISBN 5-03-002419-0
- Ivanov V. G., Berestovski T. N. biologisten kalvojen lipidikaksoiskerros. - M .: Nauka, 1982.
- Rubin A. B. Biofysiikka, oppikirja 2 osassa. - 3. painos, tarkistettu ja laajennettu. - M .: Moscow University Press, 2004. -
Eläinsolujen ulompi solukalvo (plasmalemma, sytolemma, plasmakalvo). peitetty ulkopuolelta (eli puolelta, joka ei ole kosketuksissa sytoplasmaan) kerroksella oligosakkaridiketjuja, jotka ovat kovalenttisesti kiinnittyneet kalvoproteiineihin (glykoproteiineihin) ja vähemmässä määrin lipideihin (glykolipideihin). Tätä kalvon hiilihydraattipäällystettä kutsutaan glykokaliksi. Glykokaliksin tarkoitus ei ole vielä kovin selvä; oletetaan, että tämä rakenne osallistuu solujen välisen tunnistusprosesseihin.
Kasvisoluissa ulomman solukalvon päällä on tiivis selluloosakerros, jossa on huokoset, joiden kautta kommunikointi tapahtuu naapurisolujen välillä sytoplasmisten siltojen kautta.
Solut sieniä plasmalemman päällä - tiheä kerros kitiini.
klo bakteerit – mureina.
Biologisten kalvojen ominaisuudet
1. Kyky koota itsekseen tuhoisan vaikutuksen jälkeen. Tämän ominaisuuden määräävät fosfolipidimolekyylien fysikaalis-kemialliset ominaisuudet, jotka vesiliuoksessa yhdistyvät niin, että molekyylien hydrofiiliset päät kääntyvät ulospäin ja hydrofobiset päät sisäänpäin. Proteiineja voidaan sisällyttää valmiisiin fosfolipidikerroksiin. Kyky kokoontua itse on välttämätöntä solutasolla.
2. Puoliläpäisevyys(selektiivisyys ionien ja molekyylien siirrossa). Varmistaa ioni- ja molekyylikoostumuksen pysyvyyden säilymisen solussa.
3. Kalvon juoksevuus. Kalvot eivät ole jäykkiä rakenteita, vaan ne vaihtelevat jatkuvasti lipidi- ja proteiinimolekyylien pyörimis- ja värähtelyliikkeiden vuoksi. Tämä saa aikaan suuren määrän entsymaattisia ja muita kemiallisia prosesseja kalvoissa.
4. Kalvojen fragmenteilla ei ole vapaita päitä, koska ne ovat suljettuja kupliin.
Ulomman solukalvon (plasmalemma) toiminnot
Plasmalemman päätehtävät ovat seuraavat: 1) este, 2) reseptori, 3) vaihto, 4) kuljetus.
1. estetoiminto. Se ilmenee siinä, että plasmalemma rajoittaa solun sisältöä erottaen sen ulkoisesta ympäristöstä ja solunsisäiset kalvot jakavat sytoplasman erillisiksi reaktiosoluiksi. osastoja.
2. reseptorin toiminta. Yksi plasmalemman tärkeimmistä tehtävistä on varmistaa solun kommunikaatio (yhteys) ulkoisen ympäristön kanssa kalvoissa olevan reseptorilaitteen kautta, jolla on proteiini- tai glykoproteiiniluonne. Plasmalemman reseptorimuodostelmien päätehtävä on ulkoisten signaalien tunnistaminen, minkä ansiosta solut ovat oikein suuntautuneet ja muodostavat kudoksia erilaistumisprosessissa. Erilaisten säätelyjärjestelmien aktiivisuus sekä immuunivasteen muodostuminen liittyy reseptorin toimintaan.
vaihtotoiminto määräytyy entsyymiproteiinien sisällöstä biologisissa kalvoissa, jotka ovat biologisia katalyyttejä. Niiden aktiivisuus vaihtelee riippuen väliaineen pH:sta, lämpötilasta, paineesta, sekä substraatin että itse entsyymin pitoisuudesta. Entsyymit määräävät avainreaktioiden voimakkuuden aineenvaihduntaa sekä suuntautuminen.
Kalvojen kuljetustoiminto. Kalvo mahdollistaa erilaisten kemikaalien selektiivisen tunkeutumisen soluun ja solusta ympäristöön. Aineiden kuljetus on välttämätöntä sopivan pH:n ylläpitämiseksi solussa, oikeanlaisen ionipitoisuuden ylläpitämiseksi, mikä varmistaa solun entsyymien tehokkuuden. Kuljetus toimittaa ravinteita, jotka toimivat energianlähteenä, sekä materiaalia erilaisten solukomponenttien muodostumiseen. Se määrää myrkyllisten jätteiden poistumisen solusta, erilaisten hyödyllisten aineiden erittymisen sekä hermo- ja lihastoiminnan kannalta välttämättömien ionigradienttien muodostumisen Muutokset aineiden siirtymisnopeudessa voivat johtaa häiriintyneisiin bioenergeettisissä prosesseissa, vesi-suola-aineenvaihdunnassa , kiihtyvyys ja muut prosessit. Näiden muutosten korjaaminen on monien lääkkeiden toiminnan taustalla.
On kaksi päätapaa, joilla aineet tulevat soluun ja ulos solusta ulkoiseen ympäristöön;
passiivinen kuljetus,
aktiivinen kuljetus.
Passiivinen kuljetus kulkee kemiallisen tai sähkökemiallisen pitoisuuden gradienttia pitkin kuluttamatta ATP-energiaa. Jos kuljetettavan aineen molekyylissä ei ole varausta, passiivisen kuljetuksen suunta määräytyy vain tämän aineen pitoisuuden eron perusteella kalvon molemmilla puolilla (kemiallinen pitoisuusgradientti). Jos molekyyli on varautunut, niin sen kuljetukseen vaikuttavat sekä kemiallinen pitoisuusgradientti että sähköinen gradientti (kalvopotentiaali).
Molemmat gradientit yhdessä muodostavat sähkökemiallisen gradientin. Aineiden passiivinen kuljetus voidaan suorittaa kahdella tavalla: yksinkertainen diffuusio ja helpotettu diffuusio.
Yksinkertaisella diffuusiolla suola-ionit ja vesi voivat tunkeutua selektiivisten kanavien läpi. Nämä kanavat muodostavat jotkut transmembraaniproteiinit, jotka muodostavat päästä päähän -kuljetusreittejä, jotka ovat avoinna pysyvästi tai vain lyhyen aikaa. Selektiivisten kanavien kautta tunkeutuu erilaisia molekyylejä, joiden koko ja varaus vastaavat kanavia.
Yksinkertaiseen diffuusioon on toinenkin tapa - tämä on aineiden diffuusio lipidikaksoiskerroksen läpi, jonka läpi rasvaliukoiset aineet ja vesi kulkevat helposti. Lipidikaksoiskerros ei läpäise varautuneita molekyylejä (ioneja), ja samalla varauksettomat pienet molekyylit voivat diffuusoitua vapaasti, ja mitä pienempi molekyyli, sitä nopeammin se kulkeutuu. Melko korkea veden diffuusionopeus lipidikaksoiskerroksen läpi johtuu juuri sen molekyylien pienestä koosta ja varauksen puuttumisesta.
Helpotettu diffuusio proteiinit ovat mukana aineiden kuljettamisessa - kantajia, jotka toimivat "ping-pongin" periaatteella. Tässä tapauksessa proteiini esiintyy kahdessa konformaatiotilassa: "pong"-tilassa kuljetetun aineen sitoutumiskohdat ovat avoimia kaksoiskerroksen ulkopuolella ja "ping"-tilassa samat kohdat avautuvat toisella puolella. puolella. Tämä prosessi on palautuva. Se, miltä puolelta aineen sitoutumiskohta on auki tietyllä hetkellä, riippuu tämän aineen pitoisuusgradientista.
Tällä tavalla sokerit ja aminohapot kulkevat kalvon läpi.
Helpotetun diffuusion avulla aineiden kulkunopeus kasvaa merkittävästi verrattuna yksinkertaiseen diffuusioon.
Kantajaproteiinien lisäksi jotkin antibiootit, kuten gramidiini ja valinomysiini, osallistuvat helpotettuun diffuusioon.
Koska ne tarjoavat ionien kuljetuksen, niitä kutsutaan ionoforit.
Aktiivinen aineiden kuljetus solussa. Tämäntyyppiseen kuljetukseen liittyy aina energiakustannuksia. Aktiiviseen kuljetukseen tarvittava energialähde on ATP. Tämän tyyppisen kuljetuksen tyypillinen piirre on, että se suoritetaan kahdella tavalla:
ATPaasi-nimien entsyymien avulla;
kuljetus kalvopakkauksessa (endosytoosi).
V ulompi solukalvo sisältää entsyymiproteiineja, kuten ATPaaseja, joiden tehtävänä on tarjota aktiivista kuljetusta ioneja pitoisuusgradienttia vastaan. Koska ne kuljettavat ioneja, tätä prosessia kutsutaan ionipumpuksi.
Eläinsoluissa on neljä pääasiallista ioninkuljetusjärjestelmää. Niistä kolme mahdollistaa siirtymisen biologisten kalvojen läpi: Na + ja K +, Ca +, H + ja neljäs - protonien siirto mitokondrioiden hengitysketjun toiminnan aikana.
Esimerkki aktiivisesta ioninsiirtomekanismista on natrium-kaliumpumppu eläinsoluissa. Se ylläpitää solussa vakiona natrium- ja kalium-ionien pitoisuutta, joka eroaa näiden aineiden pitoisuudesta ympäristössä: normaalisti solussa on vähemmän natriumioneja kuin ympäristössä ja enemmän kaliumia.
Tämän seurauksena yksinkertaisen diffuusion lakien mukaan kaliumilla on taipumus poistua solusta ja natrium diffundoituu soluun. Toisin kuin yksinkertainen diffuusio, natrium-kaliumpumppu pumppaa jatkuvasti natriumia ulos solusta ja ruiskuttaa kaliumia: kolmea natriummolekyyliä kohti, jotka heitetään ulos, soluun syötetään kaksi kaliummolekyyliä.
Tämän natrium-kalium-ionien kuljetuksen varmistaa ATP-riippuvainen entsyymi, joka on lokalisoitunut kalvoon siten, että se tunkeutuu sen koko paksuuteen.Natrium ja ATP tulevat tähän entsyymiin kalvon sisältä ja kalium kalvon sisältä. ulkopuolella.
Natriumin ja kaliumin siirtyminen kalvon läpi tapahtuu konformaatiomuutosten seurauksena, jotka natrium-kalium-riippuvainen ATPaasi käy läpi, ja joka aktivoituu, kun natriumin pitoisuus solun sisällä tai kaliumissa ympäristössä kasvaa.
Tämän pumpun käyttö edellyttää ATP-hydrolyysiä. Tämän prosessin tarjoaa sama natrium-kalium-riippuvainen ATP-aasi-entsyymi. Samaan aikaan yli kolmasosa eläinsolun levossa kuluttamasta ATP:stä kuluu natrium-kaliumpumpun työhön.
Natrium-kaliumpumpun moitteettoman toiminnan rikkominen johtaa erilaisiin vakaviin sairauksiin.
Tämän pumpun hyötysuhde ylittää 50%, mitä ei saavuteta edistyneimmillä ihmisen luomilla koneilla.
Monia aktiivisia kuljetusjärjestelmiä ohjaa ionigradientteihin varastoitunut energia eikä ATP:n suora hydrolyysi. Kaikki ne toimivat yhteiskuljetusjärjestelminä (helpoten pienmolekyylipainoisten yhdisteiden kuljetusta). Esimerkiksi tiettyjen sokereiden ja aminohappojen aktiivinen kuljetus eläinsoluihin määräytyy natriumionigradientin mukaan, ja mitä korkeampi natriumionigradientti on, sitä suurempi on glukoosin absorptionopeus. Päinvastoin, jos natriumin pitoisuus solujen välisessä tilassa laskee huomattavasti, glukoosin kuljetus pysähtyy. Tässä tapauksessa natriumin tulee liittyä natriumista riippuvaiseen glukoosin kantajaproteiiniin, jolla on kaksi sitoutumiskohtaa: yksi glukoosille ja toinen natriumille. Soluun tunkeutuvat natriumionit edistävät kantajaproteiinin kulkeutumista soluun glukoosin mukana. Glukoosin mukana soluun tulleet natriumionit pumpataan takaisin natrium-kaliumriippuvaisen ATPaasin toimesta, joka ylläpitämällä natriumkonsentraatiogradienttia säätelee epäsuorasti glukoosin kuljetusta.
Aineiden kuljetus kalvopakkauksissa. Suuret biopolymeerimolekyylit eivät käytännössä pysty tunkeutumaan plasmalemman läpi millään edellä kuvatuista aineiden soluunkuljetusmekanismeista. Solu sieppaa ne ja imeytyy kalvopakkaukseen, jota kutsutaan endosytoosi. Jälkimmäinen jaetaan muodollisesti fagosytoosiin ja pinosytoosiin. Solu sitoo kiinteitä hiukkasia fagosytoosi ja nestettä - pinosytoosi. Endosytoosin aikana havaitaan seuraavat vaiheet:
imeytyneen aineen vastaanotto solukalvon reseptoreiden takia;
kalvon tunkeutuminen kuplan (rakkuloiden) muodostumiseen;
endosyyttisen rakkulan erottaminen kalvosta energiankulutuksella - fagosomien muodostuminen ja kalvon eheyden palauttaminen;
Fagosomin fuusio lysosomin kanssa ja muodostuminen fagolysosomit (ruoansulatusvakuoli), jossa imeytyneiden hiukkasten pilkkominen tapahtuu;
hajoamattoman materiaalin poistaminen fagolysosomista solusta ( eksosytoosi).
Eläinten maailmassa endosytoosi on tyypillinen tapa ruokkia monia yksisoluisia organismeja (esimerkiksi ameeboissa), ja monisoluisten organismien joukossa tämän tyyppinen ruokahiukkasten pilkkominen esiintyy endodermaalisissa soluissa coelenteraateissa. Mitä tulee nisäkkäisiin ja ihmisiin, niillä on retikulo-histio-endoteliaalinen solujärjestelmä, joka kykenee endosytoosiin. Esimerkkejä ovat veren leukosyytit ja maksan Kupffer-solut. Jälkimmäiset reunustavat maksan niin kutsuttuja sinimuotoisia kapillaareja ja vangitsevat erilaisia veressä suspendoituneita vieraita hiukkasia. Eksosytoosi- tämä on myös tapa poistaa monisoluisen organismin solusta sen erittämä substraatti, joka on välttämätön muiden solujen, kudosten ja elinten toiminnalle.

